
1. Potential UHI effect on trees in the urban landscape.
Keep a cool head when selecting trees for heat tolerance
Glynn Percival and Christopher Percival
The frequency and intensity of heatwaves have increased globally over the past 20 years. Climatic modelling predicts that this trend is likely to continue throughout the 21st century with the global land area which experiences heatwaves in 2040 expected to be double that affected in 2020.
While trees with tolerance to urban stressors such as chemical pollutants, de-icing salts, drought, soil compaction and waterlogging have been identified, limited information exists regarding the tolerance of trees to prolonged heat. This article summarises what we currently know about the impact of heatwaves on trees, as well as ongoing research at the Bartlett Tree Research and Diagnostic Laboratory to identify which species offer potential for future plantings.
Introduction
Extreme temperatures associated with prolonged heatwaves, lasting for several weeks, now impact approximately 10% of land surfaces, with increases in the frequency and intensity of heatwaves world-wide expected to continue through the 21st century. Della-Marta et al. (2007) reported that the length of summer heatwaves in Western Europe has doubled and the frequency of hot days has tripled since 1880. During 2021, all-time maximum temperatures were recorded in seven US states (CA, AZ, NM, UT, CO, WY, MT). In Phoenix, Arizona, high temperatures ranged between 46 and 47°C. Similar unprecedented heatwaves were also recorded in Canada where temperatures exceed 47°C. Within the UK, the Met Office issued its first ever extreme heat warning. In conclusion, all climatic predictions state that global warming will be represented by more frequent and intense high-temperature events.
Consequently, as extreme heat events increase in frequency, it is important to select and plant trees that will be able to cope. Such a selection process is even more important within urban landscapes due to the urban heat island effect (UHI effect) where the air temperature can be 2–3°C higher than that of the surrounding rural area (Photograph 1). The UHI effect is caused by many factors including absorption of short-wave radiation from the sun by buildings and street surfaces, as well as air pollutants caused by traffic and building temperature-regulating (heating/cooling) machinery that absorb and re-emit longwave radiation into the urban environment.
Heat stress and influences on trees
Heat stress is defined as the rise in temperature beyond a threshold level for a period of time sufficient to cause irreversible damage to plant growth and development. Heat stress impacts plant function from the cellular to whole-plant scale, including damage to leaf photosynthesis, enzyme activity and leaf structure. Heat stress also increases respiration and reduces photosynthesis, stomatal conductance, growth and reproduction, leading to leaf abscission, visible foliar damage and mortality. At very high temperatures, severe cellular injury and cell death can occur within minutes (Photograph 2). In Europe, forest mortality due to drought and heat stress occurred across many Mediterranean regions throughout the 1990s and 2000s. Symptomology across forest stands included foliage yellowing and browning, branch dieback, premature leaf loss and tree death. Oak, fir, spruce, beech and pine have been severely affected, raising concerns about the consequences for forest health. Similar scales of tree mortality have been recorded in the US, Asia, Australasia and South and Central America following heat and drought stress episodes, raising concern about how prolonged heatwaves have the potential to alter forest structure, composition and subsequent ecosystem services such as carbon sequestration.
Temperature tolerance
The temperature optimum for boreal, temperate and tropical angiosperm and gymnosperm trees ranges between 20 and 30°C. However, many species can tolerate temperatures up to and between 35 and 40°C without any major detrimental effects on their biology. A number of tropical tree species adapted to thrive in high-temperature climates, e.g. Dipteryx oleifera and Zygia longifolia, can maintain positive net photosynthesis at temperatures ≥45°C. Damage to leaf photosynthetic properties caused by temperatures below 40°C is generally reversible irrespective of tree species. However, prolonged temperatures ≥40°C tend to cause irreversible damage, although effects can be influenced by species, duration of temperature stress and prior temperature exposure (Yordanov, 1992). Experiments using seedlings of seven common UK tree species (Acer pseudoplatanus, Betula pendula, Fagus sylvatica, Fraxinus excelsior, Juglans regia, Quercus petraea, Q. robur) grown under nursery conditions revealed that the critical temperature at which heat damage occurred was around 47°C, with no significant difference among species (Dreyer et al., 2001). However, based on further work evaluating the heat tolerance of several temperate and boreal trees (Quercus muehlenbergii, Q. macrocarpa, Q. suber, Q. canariensis, Q. robur, Q. petraea, Picea glauca, Populus deltoides, Ficus insipida, Acer pseudoplatanus, Betula verrucosa, Fagus sylvatica, Fraxinus excelsior, Juglans regia), a value of ≥42°C is proposed as a critical temperature from which trees suffer irreversible damage rather than 47°C. Consequently, it is suggested that ≥42°C be used as a guide for urban landscape managers and arborists (Teskey et al., 2015).
Allen et al. (2010) importantly emphasises the fact that many trees die not directly from heat-and drought-induced episodes but indirectly by insect and diseases ingress caused by repeat years of drought and heat stress which weaken them. Within the UK, for example, changes in drought/heat intensity and frequency have potentially altered the impacts, populations and severity of tree-damaging insect and fungal pathogens. Armillaria, Heterobasidion irregulare and H. occidentale, cankers (Septoria musiva, Diplodia sapinea, Hymenoscyphus fraxineus, Cryphonectria parasitica, Massaria [Splanchnonema platani]) and bark-boring beetles such as Agrilus or Scolytus are associated with increased drought and heat episodes (Percival and Banks, 2019).
Tree selection: heat and/or drought?
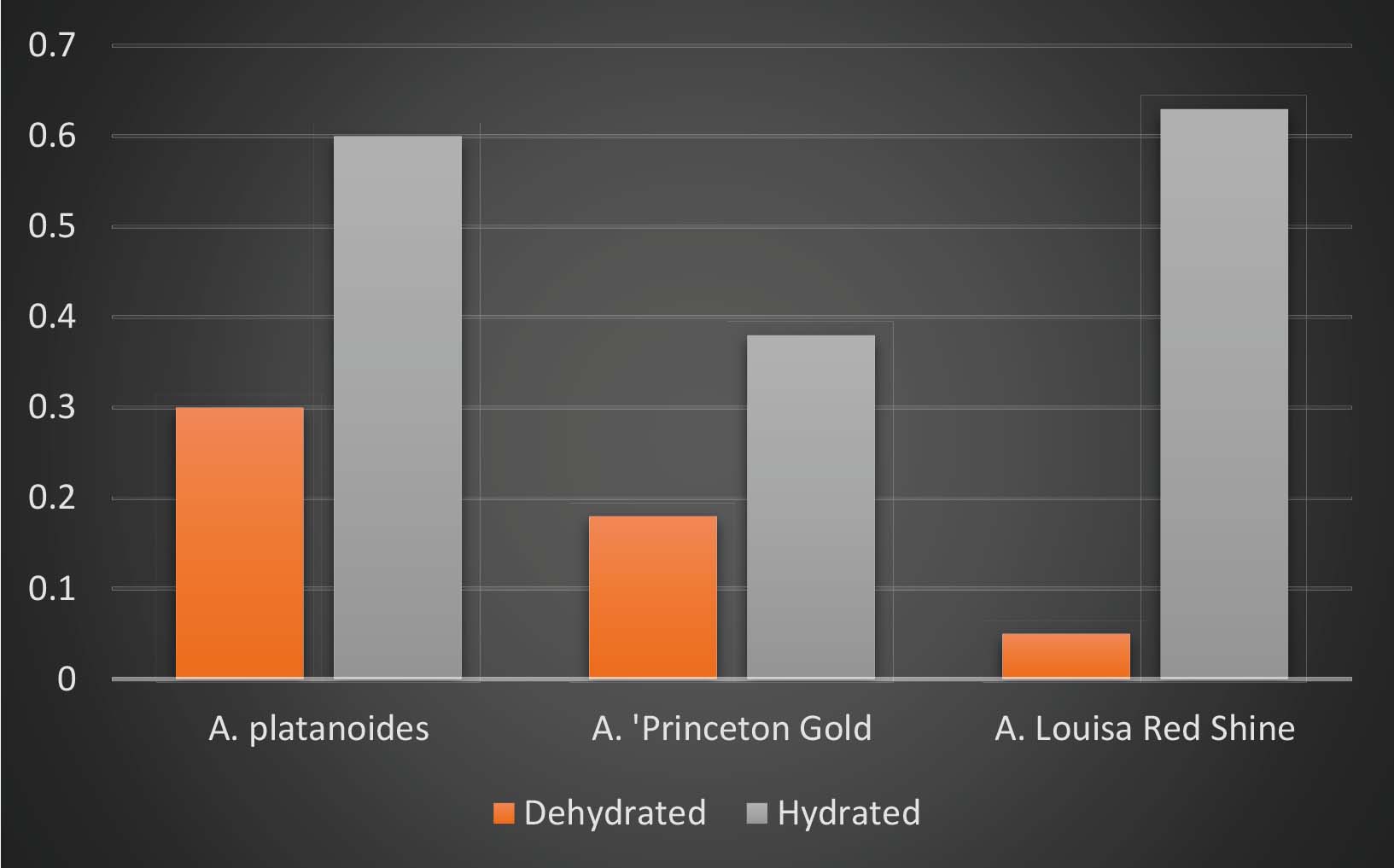
Figure 1: The influence of heat stress on leaf photosynthetic properties of three Acer species as measured by chlorophyll fluorescence Fv/Fm values.
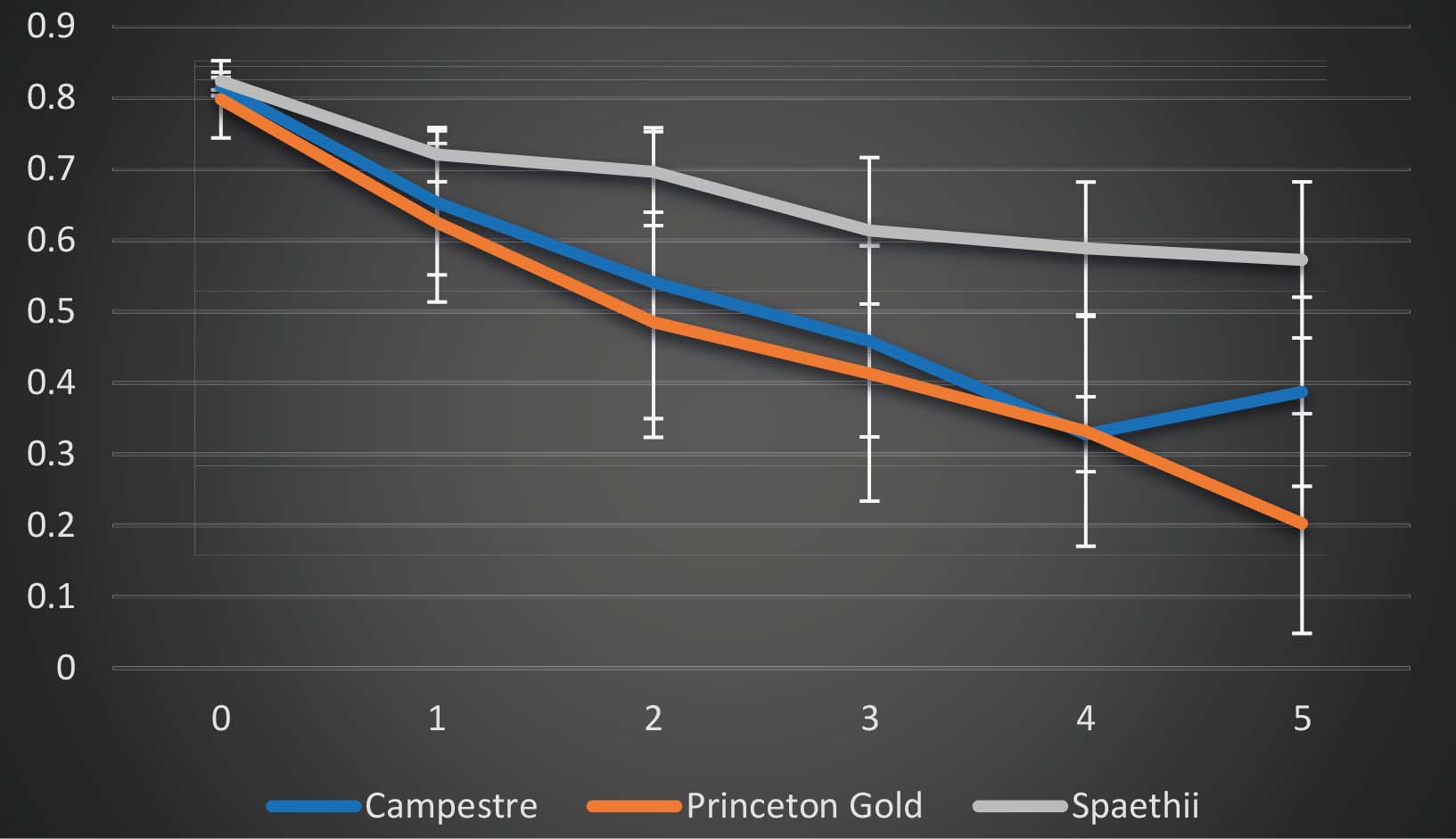
Figure 2: The influence of 5 hours at 44oC (x-axis) on leaf photosynthetic properties of three Acer species as measured by chlorophyll fluorescence Fv/Fm values (y-axis).
One of the great challenges for urban landscape managers will be to select heat-tolerant trees for future predicted climatic conditions. These include extreme heat events coupled with increased frequency and duration of summer heatwaves, as well as increased monthly maximum temperatures during the growing season.
As heat and drought are often concurrent stress factors in nature then, by default, selection for drought resistance is also associated with selection for heat tolerance as both stressors tend to be intrinsically linked. Heat stress, for example, under conditions of high soil moisture is relatively rare. However, the assumption may be flawed as the two stressors exert very different effects on, for example, leaf photosynthetic processes. The primary target of heat is the thylakoid membrane, in particular impairment of photosystem II (Schrader et al., 2004). Drought, on the other hand, does not directly impair the bio- and photochemistry of photosynthesis, but it primarily down-regulates photosynthesis by increasing diffusive resistances to CO2 entry into the chloroplasts.
In a detailed study, Correia et al. (2018) quantified physiological, biochemical and metabolomic changes in a drought-tolerant Eucalyptus globulus clone in response to drought and heat stress that were imposed separately. Drought stress induced very different changes to plant physiology compared to heat stress. In response to drought, reduced pre-dawn water potential (ψpd) and gas exchange were accompanied by increased malondialdehyde and a total glutathione leaf content, indicative of oxidative stress. Abscisic acid (ABA) levels increased while jasmonic acid (JA) content fell. Metabolic alterations included reduced levels of sugar phosphates accompanied by increased starch and non-structural carbohydrates.
On the other hand, heat stress triggered an increase in relative water content (RWC) and ψpd. Photosynthetic rate and leaf chlorophyll pigments were reduced, and this was accompanied by a reduction in water-use efficiency, salicylic acid and JA content. Sugar alcohols and several amino acids were enhanced, while starch, fructose-6-phosphate, glucose-6-phosphate and α-glycerophosphate were reduced.
Little, if any, research exists showing a clear association between heat and drought stress in trees, i.e. that selection of drought-tolerant trees equates to heat-tolerant trees. With respect to crops, where far more research exists, results are conflicting. A clear relationship between drought and heat tolerance has been shown in chickpea and sorghum. However, with respect to rice and peanut, drought tolerance does not necessarily confer heat tolerance; indeed, many drought-tolerant cultivars have been shown to be highly sensitive to heat stress (Wahid et al., 2007).
As a precursor to larger studies, research at the Bartlett Tree Research and Diagnostic Laboratory subjected leaves of three species of Acer to heat stress under laboratory conditions. Half of the leaves were kept hydrated during the heat stress imposed (heat only) and half dehydrated to simulate both heat and drought. At the end of the experiment the influence of heat on leaf chlorophyll fluorescence Fv/Fm responses was recorded as a measure of damage to the leaf photosynthetic system. Results (Figure 1) clearly show that leaves which were kept hydrated throughout the heat stress had 50–90% less damage to their photosynthetic system, emphasising the importance of irrigation throughout a heatwave. In addition, results also showed a marked difference in heat tolerance between species with A. campestre ‘Louisa Red Shine’ and A. platanoides having a higher heat tolerance than A. ‘Princeton Gold’.
It is well recognised that maintaining a constant water status is a critical variable during heat stress (Hassan et al., 2020). Heat stress, however, can severely impair this tendency when water is limited. Under these conditions stomata close to reduce water loss. A significant survival strategy in response to heat stress is that stomata remain open during prolonged elevated temperature episodes as transpirational cooling of the leaf minimises heat damage (Ameye et al., 2012).
Tree selection for heat tolerance based on scientific quantification

2. Symptoms of prolonged heat stress on rhododendron.
Although there are EU and US guides for tree selection for urban landscapes that identify drought-, waterlogging- and shade-tolerant species, no criteria exist for heat tolerance (citree.de/about.php?language=en; www.arborday.org/trees/index-choosing.cfm).
Likewise, in relation to urban tree species little study has taken place with respect to heat adaptation and/or subsequent tolerance. Based on the research that is available, Pinus taeda and Quercus rubra seedlings exposed to repeated moderate (+6°C) or extreme (+12°C) heatwaves under greenhouse conditions resulted in significant growth reductions at +12°C, but not +6°C (Bauweraerts et al., 2014). Cedrus brevifolia was shown to be more heat tolerant than C. libani and C. atlantica when seedlings of the three species were exposed to temperatures above 45°C for 5 hours (Ladjal et al., 2000), while populations of Pseudotsuga menziesii were shown to be more heat tolerant than populations of Pinus ponderosa by Marias (2017). Urban landscapers, planners and arborists are faced with thousands of ornamental tree species to select for urban landscape plantings, yet the bulk of scientific research into quantification of heat tolerance has focused on a limited number of genera (Quercus, Pinus, Eucalyptus, Acer, Picea) and within those genera only a limited number of species, which in turn are predominately planted for forestry purposes.
Recent research at the Bartlett Tree Research and Diagnostic Laboratory has identified variation in the heat-stress response of three Acer species (Figure 2). In this experiment leaves of A. campestre, A. ‘Princeton Gold’ and A. spaethii were subjected to 5 hours at 44°C and the effects on leaf chlorophyll fluorescence Fv/Fm responses recorded hourly as a measure of damage to the leaf photosynthetic system. Results clearly show a species-specific response, with heat tolerance in the order A. spaethii>A. campestre>A. ‘Princeton Gold’. Further studies are ongoing.
Tree selection based on experience
A number of heat-tolerant urban tree species have been identified and/or recommended by organisations such as the Royal Horticultural Society, UK (www.rhs.org.uk/advice/profile?PID=712), or US university-based extension services (extension.psu.edu/heat-and-drought-tolerant-plants). These selection processes appear to be based on, or restricted to, either post-event heat analysis and/or observation of species that grow well locally in hot ‘site locations’ – i.e. close to large masses of asphalt and concrete, close to and adjacent to buildings, near underground utilities and/or in containers/raised beds that were recorded as hotter than in-ground planting areas – rather than on actual scientific methodology. For example, Table 1 is amended from Appleton et al. (2015), recommending heat-tolerant tree species for urban landscape plantings.
Table 1: Trees for hot sites.
Common name |
Latin name |
Field maple |
Acer campestre |
Red maple |
Acer rubrum |
Sugar maple |
Acer saccharum |
Freeman maple |
Acer × freemanii |
Red horse chestnut |
Aesculus × carnea |
River birch |
Betula nigra |
White birch |
Betula platyphylla |
Cockspur hawthorn |
Crataegus crus-galli |
Japanese cryptomeria |
Cryptomeria japonica |
Leyland cypress |
× Cupressocyparis leylandii |
Ginkgo |
Ginkgo biloba |
Honey locust |
Gleditsia triacanthos |
Chinese juniper |
Juniperus chinensis |
Sweetgum |
Liquidambar styraciflua |
Colorado spruce |
Picea pungens |
London plane |
Platanus × acerifolia |
Pissard plum |
Prunus cerasifera |
Yoshino cherry |
Prunus × yedoensis |
Callery pear (cultivars) |
Pyrus calleryana |
English oak |
Quercus robur |
Red oak |
Quercus rubra |
Japanese pagoda tree |
Sophora japonica |
White cedar |
Thuja occidentalis |
Oriental arborvitae |
Thuja orientalis |
Small leaf lime |
Tilia cordata |
Silver lime |
Tilia tomentosa |
Tree selection based on provenance
Provenance selection also offers an abundance of largely untapped genetic resource to select for heat-tolerant trees. Robakowski et al. (2012) investigated the temperature responses of Acer rubrum and Quercus rubra. The provenances of A. rubrum and Q. rubra that originated from northern sites with a lower ambient temperature had a lower photosynthetic temperature optimum compared to A. rubrum and Q. rubra that originated from southern provenances. Weston and Bauerle (2007) demonstrated that variation exists in the response of A. rubrum to excessive heat. Trees collected from Florida (yearly average temperature of 22.6°C) maintained close to a two-fold increase in leaf photosynthetic rates and had a 22% increase in the maximal rate of RUBISCO (ribulose bisphosphate carboxylase–oxygenase) carboxylation at temperatures of 33–42°C compared to trees from Minnesota (yearly average temperature of 3°C). Research introducing Acer saccharum of three Canadian provenances with mean annual temperatures of 3°C (Manitoba), 4.2°C (Quebec) and 9.4°C (Ontario) to a site in subtropical China (15.8°C) monitored survival, growth and summer photosynthetic rates under field and growth chamber conditions. The Ontario provenance had the highest propensity for survival and growth, followed by the Quebec provenance, while the Manitoba provenance had the lowest. Likewise, the Ontario provenance had a higher photosynthesis rate and stomatal conductance than the Quebec and Manitoba provenances. Overall, the Ontario provenance had the best physiological adjustment for self-protection from heat stress, followed by the Quebec and then the Manitoba provenances (Zhu et al., 2019). Further work supporting the potential of provenance selection for heat tolerance in Eucalyptus globulus and E. pauciflora is provided by Pita et al. (2005) and Ferrar et al. (1989). For future research purposes identification of species from the warmest portion of their range may be critical in selection of heat-tolerant trees.
Conclusions
Selection of heat-tolerant trees will become of greater importance as the warming effects of climate change increase the intensity of the UHI effect within UK urban landscapes. This in turn will increase levels of indoor and outdoor thermal discomfort, potentially inducing adverse health effects such as sunburn, skin cancer and cataracts. Presently little information exists regarding the heat tolerance of urban trees. The information that is available recognises that trees possess physiological and biochemical mechanisms to allow them to survive extreme heat events. These present opportunities for research that will identify superior heat-tolerant trees as well as increase our knowledge of how urban trees acclimate and adapt to heat stress. Understanding the mechanisms of tree responses to extreme heat temperature events will also be critically important for understanding how tree species will be affected by climate change.
References used to support this article
Allen, C.D., Macalady, A.K., Chenchouni, H., Bachelet, D., McDowell, N., Vennetier, M., Cobb, N. (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management 259: 660–684.
Ameye, M., Wertin, T.M., Bauweraerts, I., McGuire, M.A., Teskey, R.O., Steppe, K. (2012). The effect of induced heat waves on Pinus taeda and Quercus rubra seedlings in ambient and elevated CO2 atmospheres. The New Phytologist 196: 448–461.
Appleton, B., Rudiger, E.L.T., Harris, R., Sevebeck, K., Alleman, D., Swanson, L. (2015). Trees for Hot Sites. Virginia Cooperate Extension, Virginia Tech, Virginia State University Technical Note Publication 430-024.
Bauweraerts, I., Ameye, M., Wertin, T.M., McGuire, M.A., Teskey, R.O., Steppe, K. (2014). Water availability is the decisive factor for the growth of two tree species in the occurrence of consecutive heat waves. Agricultural and Forest Meteorology 189–190: 19–29.
Correia, B., Hancock, R.D., Amaral, J., Gomez-Cadenas, A., Valledor, L., Pinto, G., et al. (2018). Combined drought and heat activates protective responses in Eucalyptus globulus that are not activated when subjected to drought or heat stress alone. Frontiers Plant Science doi.org/10.3389/fpls.2018.00819
Della-Marta, P.M., Haylock, M.R., Luterbacher, J., Wanner, H. (2007). Doubled length of western European summer heat waves since 1880. Journal of Geophysical Research-Atmospheres 112: D15103.
Dreyer, E., Le Roux, X., Montpied, P., Daudet, F.A., Masson, F. (2001). Temperature response of leaf photosynthetic capacity in seedlings from seven temperate tree species. Tree Physiology 21: 223–232.
Ferrar, P.J., Slatyer, R.O and Vranjic, J.A. (1989). Photosynthetic temperature acclimation in Eucalyptus species from diverse habitats, and a comparison with Nerium oleander. Australian Journal of Plant Physiology 16(2): 199–217.
Hassan, M.U., Chattha, M.U., Khan, I., Chattha, M.B., Barbanti, L., Aamer, M., Iqbal, M.M., Nawaz, M., Mahmood, A., Ali., A, Aslam, M.T. (2020). Heat stress in cultivated plants: nature, impact, mechanisms, and mitigation strategies – a review. Plant Biosystems 155(7): 1–56.
Ladjal, M., Epron, D., Ducrey, M. (2000). Effects of drought preconditioning on thermo-tolerance of photosystem II and susceptibility of photosynthesis to heat stress in cedar seedlings. Tree Physiology 20(18): 1235–1241.
Marias, D.E., Meinzer, F.C., Woodruff, D.R., McCilloh, K.A. (2017). Thermo-tolerance and heat stress responses of Douglas-fir and ponderosa pine seedling populations from contrasting climates. Tree Physiology 37(3): 301–315.
Percival, G.C, Banks J.M. (2019). Drought stress effects on pests and diseases. ARB Magazine 186: 68–71.
Pita P., Cañas I., Soria F., Ruiz F., Toval G. (2005). Use of physiological traits in tree breeding for improved yield in drought-prone environments. The case of Eucalyptus globulus. Investigación Agraria. Sistemas y Recursos Forestales 14: 383–393.
Robakowski, P., Li, Y., Reich, P.B. (2012). Local ecotypic and species range-related adaptation influence photosynthetic temperature optima in deciduous broadleaved trees. Plant Ecology 213(1): 113–125.
Schräder, S.M, Wise, R.R., Wacholtz, W.F., Ort, D.R., Sharkey, T.D. (2004). Thylakoid membrane responses to moderately high leaf temperature in Pima cotton. Plant, Cell & Environment 27, 725-735.
Teskey, R., Wertin, T., Bauweraerts I., Ameye, M., McGuire, M.A and Steppe, K. (2015). Responses of tree species to heat waves and extreme heat events. Plant, Cell & Environment 38: 1699–1712.
Wahid A., Gelani S., Ashraf M., Foolad, M.R. (2007). Heat tolerance in plants: an overview. Environmental and Experimental Botany 61: 199–223.
Weston, D.J., Bauerle, W.L. (2007). Inhibition and acclimation of C3 photosynthesis to moderate heat: a perspective from thermally contrasting genotypes of Acer rubrum (red maple). Tree Physiology 27: 1083–1092.
Yordanov, I. (1992). Response of photosynthetic apparatus to temperature stress and molecular mechanisms of its adaptations. Photosynthetica 26: 517–531.
Zhu. Y., Fu, S., Liu, H., Wang, Z., Han, Y.H.C (2019). Heat stress tolerance determines the survival and growth of introduced Canadian sugar maple in subtropical China. Tree Physiology 1;39(3):417–426. doi.org/10.1093/treephys/tpy098.
Dr Glynn Percival is the senior arboricultural researcher at the Bartlett Tree Research and Diagnostic Laboratory at the University of Reading UK.
Christopher Percival is a research technician at the Bartlett Tree Research and Diagnostic Laboratory at the University of Reading UK.
This article was taken from Issue 197 Summer 2022 of the ARB Magazine, which is available to view free to members by simply logging in to the website and viewing your profile area.
